(Ciencias de Joseleg)(Biología)(Introducción y biología
celular)(Los
organelos celulares)(Introducción)(El
citosol)(Organelos
procariotas)(Los
retículos endoplasmáticos)(El
aparato de Golgi)(El
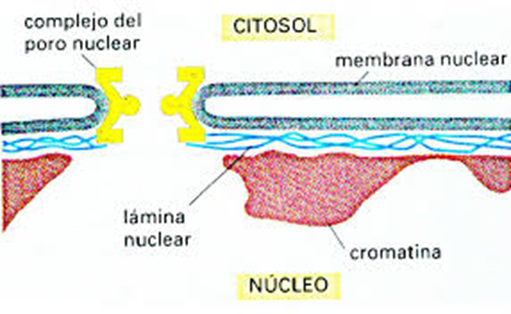
núcleo celular)(Vesículas
celulares)(La
mitocondria y el cloroplasto)(Referencias
bibliográficas)(Versión
documento word)
Alberts, B., Johnson, A., Lewis, J.,
Morgan, D., Raff, M., Roberts, K., & Walter, P. (2015). Molecular
biology of the cell (5th ed.). Garland Science.
Antonopoulos,
E., Freisleben, H.-J., Krisnamurti, D. G. B., Estuningtyas, A., Mulyanto, C.,
Ridwan, R., & Freisleben, S. K. U. (2013). Fractionation and purification of
membrane lipids from the archaeon Thermoplasma acidophilum DSM 1728/10217. Separation
and Purification Technology, 110, 119–126.
Bardy, S. L., Ng, S. Y. M., &
Jarrell, K. F. (2004). Recent advances in the structure and assembly of the
archaeal flagellum. Journal of Molecular Microbiology and Biotechnology,
7(1–2), 41–51.
Belk, C., & Maier, V. B. (2013). Biology
Science for Life with physiology. (Pearson, Ed.) (4th ed.).
Berg, J. M., Tymoczko, J. L., Gatto,
G. J., & Stryer, L. (2015). Biochemistry (8th ed.). Freeman.
Brooker, R. J., Widmaier, E. P.,
Graham, L. E., & Stiling, P. D. (2017). Biology (4th ed.).
McGraw-Hill Education.
Campbell, M. K., & Farrell, S. O.
(2009). Biochemistry (6th ed.). USA: Thomsom Brooks/Cole.
Cox, M. M., Doudna, J. A., &
O’Donnell, M. (2012). Molecular biology (1st ed.). Freeman.
Fevrier, B., & Raposo, G. (2004).
Exosomes: endosomal-derived vesicles shipping extracellular messages. Current
Opinion in Cell Biology, 16(4), 415–421.
Grenier, D., & Mayrand, D.
(1987). Functional characterization of extracellular vesicles produced by
Bacteroides gingivalis. Infection and Immunity, 55(1), 111–117.
Hoefnagels, M. (2015). Biology:
concepts and investigations (3rd ed.). McGraw-Hill New York.
Kåhrström, C. T. (2013). Biofilms:
Flagella function as mechanosensors. Nature Reviews Microbiology, 11(9),
597.
Kalanetra, K. M., Huston, S. L.,
& Nelson, D. C. (2004). Novel, attached, sulfur-oxidizing bacteria at
shallow hydrothermal vents possess vacuoles not involved in respiratory nitrate
accumulation. Applied and Environmental Microbiology, 70(12),
7487–7496.
Karp, G. C. (2013). Cell and
Molecular Biology, Concepts and Experiments (7th ed.). USA: Wiley Online
Library.
König, H. (1988). Archaeobacterial
cell envelopes. Canadian Journal of Microbiology, 34(4), 395–406.
Mackean, D. G., & Hayward, D.
(2014). Biology (3rd ed.). IGCSE Cambridge.
Mader, S. S. (2010). Biology
(10th ed.). McGraw-Hill Education.
Mader, S. S., & Windelspecht, M.
(2015). Biology (12th ed.). McGraw-Hill Education.
Mader, S. S., & Windelspecht, M.
(2018). Essentials of biology (5th ed.). McGraw-Hill Education.
Mao, Y. S., Zhang, B., & Spector,
D. L. (2011). Biogenesis and function of nuclear bodies. Trends in Genetics,
27(8), 295–306.
Mashburn‐Warren, L. M., &
Whiteley, M. (2006). Special delivery: vesicle trafficking in prokaryotes. Molecular
Microbiology, 61(4), 839–846.
Mason, K. A., Losos, J. B., Singer,
S. R., & Raven, P. H. (2014). Biology (7th ed.). McGraw-Hill New
York.
Mounkes, L. C., & Stewart, C. L.
(2004). Aging and nuclear organization: lamins and progeria. Current Opinion
in Cell Biology, 16(3), 322–327.
Murray, R. K., Bender, D. A., Botham,
K. M., Kennelly, P. J., Rodwell, V., & Weil, A. (2012). Harpers
Illustrated Biochemistry (29th ed.). McGraw-Hill Medical.
Nunes-Alves, C. (2015). Bacterial
evolution: Resurrecting motility. Nature Reviews Microbiology, 13(4),
187.
Pollard, T. D., Earnshaw, W. C.,
Lippincott-Schwartz, J., & Johnson, G. T. (2017). Cell Biology (3rd
ed.). Elsevier.
Raposo, G., & Stoorvogel, W.
(2013). Extracellular vesicles: exosomes, microvesicles, and friends. The
Journal of Cell Biology, 200(4), 373–383.
Reece, J. B., Urry, L. A., Wasserman,
S. A., Cain, M. L., Minorsky, P. V, & Jackson, R. B. (2014). Campbell
Biology (10th ed.). Pearson Higher Ed.
Sadava, D., Berenbaum, M., &
Hillis, D. (2014). Life the Science of Biology (10th ed.). Sinauer &
MacMillian.
Simon, E. J., Dickey, J. L., &
Reece, J. B. (2013). Essential biology (Pearson).
Solomon, E., Martin, C., Martin, D.
W., & Berg, L. R. (2014). Biology (10th ed.). Cengage Learning.
Starr, C., Evers, C., & Starr, L.
(2013). Biology: Today and Tomorrow With Physiology (4th ed.).
Brooks/Cole.
Taylor, M. R., Simon, E. J., Dickey,
J. L., Hogan, K., & Reece, J. B. (2018). Biology, Concepts and
conections (9th ed.). Boston: Pearson education.
Thomas, N. A., Bardy, S. L., &
Jarrell, K. F. (2001). The archaeal flagellum: a different kind of prokaryotic
motility structure. FEMS Microbiology Reviews, 25(2), 147–174.
Urry, L. A., Cain, M. L., Minorsky,
P. V, Wasserman, S. A., & Reece, J. B. (2017). Campbell Biology
(11th ed.). Pearson.
Weaver, R. F. (2012). Molecular
biology (5th ed.). McGraw-Hill Education.
Yang, L. L., & Haug, A. (1979).
Structure of membrane lipids and physico-biochemical properties of the plasma
membrane from Thermoplasma acidophilum, adapted to growth at 37 C. Biochimica
et Biophysica Acta (BBA)-Lipids and Lipid Metabolism, 573(2),
308–320.
Zhang, Z., & Zhang, R. (2015).
Epigenetics in autoimmune diseases: pathogenesis and prospects for therapy. Autoimmunity Reviews, 14(10),
854–863.















{kind=link}
{kind=link}