
(Ciencias de Joseleg)(Biología)(Introducción y biología celular)(La fotosíntesis) (Introducción)(Generalidades de la fotosíntesis)(Física de la fotosíntesis)(Destino de los átomos en la fotosíntesis)(Reacciones de la luz 1, la oxidación del agua)(Reacciones de la luz 2, Cadena de transporte de electrones y fosforilación)(Introducción a las reacciones de la oscuridad)(Etapa de fijación de carbono en las plantas C3)(Etapa de fijación de carbono en las plantas CAM y C4)(Etapa de regeneración de la RuBP)(Etapa de síntesis de carbohidratos)(Evolución de la fotosíntesis)(Referencias bibliográficas)(Versión documento word)
El ciclo de Calvin (también conocido como ciclo de Benson-Calvin) es el
conjunto de reacciones químicas que tienen lugar en los cloroplastos durante la
fotosíntesis. El ciclo es independiente de la luz porque tiene lugar después de
que la energía ha sido capturada de la luz solar. En esta sección estudiaremos
la primera mitad del ciclo, que va desde la integración de un carbono
inorgánico al sistema viviente a través de la enzima rubisco y una molécula
llamada RuBP hasta la obtención de un precursor activo de tres carbonos llamado
G3P, el cual es la base para fabricar la glucosa en reacciones independientes
al ciclo de Calvin.
La aproximación original de Calvin y colaboradores de que
una molécula de 2 carbonos era el aceptor inicial para la fijación del dióxido
de carbono resultó ser errónea, y en su lugar propusieron a una molécula de 5
carbonos llamada ribulosa-1,5-bifosfato (RuBP).
15

Fijación del carbono
en el CO2
RuBP capta un carbono del dióxido de carbono transformándose
en una molécula de 6 carbonos. El proceso es realizado por una enzima que
también participa en el segundo paso del cíclo llamada ribulosa-1,5-bifosfato
carboxilasa, conocida coloquialmente como Rubisco. La enzíma rubisco en este
paso se encarga de adicionar una molécula de dióxido de carbono a la RuBP para
formar un intermediario de 6 carbonos inestable y de muy corta duración, al
cual denominaremos RuBP+CO2. La RuBisCO que se observa en los
plastos es una proteína oligómera formada por 16 polipéptidos, que son de dos
tipos, uno grande (subunidad L) y otro pequeño (subunidad S). La subunidad L es
el sitio catalítico de esta enzima, mientras que la subunidad S es el sitio
regulador. El sitio catalítico consta de un núcleo de Mg+2 y una
lisina que tiene un CO2 unido a su extremo terminal.
El Segundo paso es el rompimiento del intermediario de 6
carbonos en dos moléculas de PGA “fosfogliceraldehído” isoméricas. Esto implica
que desde este punto se describe el ciclo en términos de una sola de estas
moléculas de tres carbonos. El hecho de que el intermediario de 6 carbonos sea
de muy corta duración explica porque los experimentos iniciales de Calvin y
colaboradores conducían a pensar que el aceptor inicial era de 2 carbonos y no
de 5 como fue determinado más tarde. Más aún, este intermediario de 6 carbonos
no se identifica en los esquemas del ciclo de Calvin que he analizado hasta el
momento. El proceso también es catalizado por la enzima rubisco. Cabe anotar
que esta enzima es de muy baja eficiencia, siendo capaz únicamente de fijar
tres moléculas de carbono por segundo, lo cuál puede ser reconocido como la
velocidad más patética de catálisis, de cualquier enzima biológica en el
planeta.
16
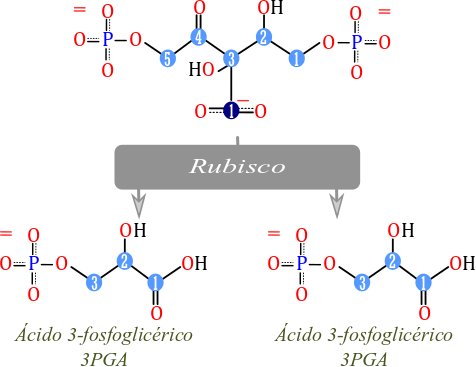
Rompimiento del intermediario de 6 carbonos a dos intermediarios de
3 carbonos.
Para compensar este defecto, prácticamente la mitad de las
proteínas que producen las células vegetales son Rubisco. De hecho, la enzima
rubisco constituye la proteína más abundante del planeta, representando entre
5-10 kg por cada ser humano.
En este paso, se produce una primera carga de energía en forma del sacrificio de un grupo fosfato del ATP a la molécula de PGA que ya tenía un grupo fosfato propio, es decir, la molécula resultante tendrá dos grupos fosfato y un mayor nivel energético. Cabe recordar que este proceso se da con cada uno de los PGA, y por cada ciclo se producen dos PGA, por lo anterior se puede decir que para fijar un solo carbono se debe sacrificar en este paso dos moléculas de ATP.
17

Carga de energía proveniente de las reacciones de la luz para
convertir el intermediario de 3 carbonos en un segundo intermediario de 3
carbonos.
La enzima que cataliza esta reacción se denomina quinasa de
fosfoglicerato, cuya función primordial es realizar una transferencia
reversible de grupos fosfato.
Esta es una segunda reacción de carga de energía, en forma
de transferencia de un par de electrones de alta energía y sus correspondientes
protones hacia el BPGA “reducción”. El resultado de la reacción se denomina
gliceraldehído 3-fosfáto “o GAP” justo con NADP. Nuevamente, por cada molécula
de dióxido de carbono que es fijada se deben sacrificar dos moléculas de NADPH
en este paso del ciclo. La enzima que cataliza esta reacción se denomina
deshidrogenasa de gliceraldehído 3-fosfáto. Un aspecto que se vuelve aparente
al analizar las enzimas y los intermediarios del ciclo de Calvin es que parecen
reciclajes de la glucolisis. Un ejemplo de esto es esta enzima, ya que esta
cataliza uno de los pasos de la glucolisis.
18

La reacción es reversible y depende de las condiciones de
equilibrio, así como la presencia de NAD/NADH “glucolisis” o NADP+/
NADPH “ciclo de Calvin”. El GAP producto de esta reacción puede ser o enviado a
la ruta de síntesis de carbohidratos, en la cual puede describirse a este grupo
de reacciones como una ruta metabólica; o, por el contrario, ser enviado a la
ruta de regeneración de RuBP la molécula de 5 carbonos empleada en el primer
paso, lo cual si completa el ciclo. Como se dijo anteriormente, por cada 12
moléculas de GAP que quedan en la bifurcación, 10 son enviadas para completar
el ciclo y generar más RuBP, y dos son enviadas por la ruta de síntesis de
carbohidratos.
El nombre es rimbombante, sin embargo, el punto es la
disponibilidad de electrones de alta energía provenientes del fotosistema I.
Anteriormente ya fue discutido que los electrones provenientes del fotosistema
I no siempre terminan en la NADP+ para formar NADPH. Los electrones
de alta energía pueden terminar en una amplia gama de aceptores finales, entre
los cuales para el presente artículo destaca una proteína denominada
tierodoxina. Tierodoxina puede ser un aceptor de electrones de alta energía, y
de hecho este evento es lo que activa o desactiva su función. Cuando la
tierodoxina recibe electrones, esta procede a reaccionar con enzimas clave del
ciclo de Calvin. Estas enzimas afectadas por la tierodoxina son activadas
iniciando el proceso de reacciones del ciclo de Calvin. Esto solo funciona
mientras que el Sol sostiene la producción de electrones de alta energía, pero
cuando llega la noche, la producción de electrones de alta energía se detiene,
y la activación de la tierodoxina se detiene. Poco tiempo después de que llega
la noche, el ciclo de Calvin se detiene debido a que la tierodoxina ya no
activa a las enzimas del ciclo. Este detalle implica que el nombre de reacciones
de la oscuridad o fase oscura de la fotosíntesis es un nombre erróneo, ya que,
de hecho, poco después de que cae la noche el ciclo de Calvin también se
detiene.




No hay comentarios:
Publicar un comentario