
(Ciencias de Joseleg)(Biología)(Introducción y biología celular)(La fotosíntesis) (Introducción)(Generalidades de la fotosíntesis)(Física de la fotosíntesis)(Destino de los átomos en la fotosíntesis)(Reacciones de la luz 1, la oxidación del agua)(Reacciones de la luz 2, Cadena de transporte de electrones y fosforilación)(Introducción a las reacciones de la oscuridad)(Etapa de fijación de carbono en las plantas C3)(Etapa de fijación de carbono en las plantas CAM y C4)(Etapa de regeneración de la RuBP)(Etapa de síntesis de carbohidratos)(Evolución de la fotosíntesis)(Referencias bibliográficas)(Versión documento word)
La radiación electromagnética es una cosa extraña a la cual los físicos describen como onda/partícula, es decir, expone características de onda, como la posesión de longitudes de onda y fenómenos como la reflexión y difracción de las ondas. Como partícula posee la capacidad para transmitir energía en unidades discretas llamadas cuantos. Dependiendo de la longitud de onda, la radiación electromagnética interactúa de forma diferencial con la materia, específicamente aquella radiación que está entre los 400 nm y los 700 nm estimula nuestros receptores visuales, y la llamamos luz, y a sus cuantos los llamamos fotones. Independientemente a su naturaleza final, a la luz en esta serie de escritos la manejamos como esa dualidad, en ocasiones la manejamos como una partícula "cuanto mencionamos los cuantos y fotones" y en otras como onda "longitudes de onda medidas en nanómetros nm".
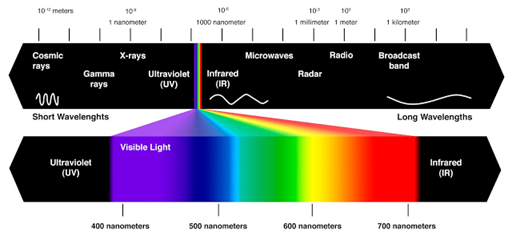
Figura 2. de onda mayor energía “violetas y ultravioletas”.
Una mol de fotones con una longitud de onda de 680 nm
contiene cerca de 45 kcal de energía, lo cual es equivalente a un cambio de
potencial redox de 1.8 voltios. La absorción de luz es el primer paso en
cualquier proceso fotoquímico. Cuando el fotón es absorbido por una molécula, uno
de sus electrones se estimula para pasar a un estado energético de mayor
energía.
Cada sustancia en la naturaleza puede absorber algunas
determinadas longitudes de onda, mientras que otras, las refleja como si fuera
un espejo, esto genera marcas distintivas de cada sustancia química en forma de
los espectros de absorción o de emisión en gases o los colores de los sólidos y
líquidos. En términos más físicos, lo que pasa es que los electrones de las
moléculas interactúan con fotones de determinadas longitudes de onda,
absorbiéndolos y aumentando su estado energético, proceso llamado
excitación. Los estados excitados de los
electrones son muy inestables y tienden a durar solo nanosegundos. Numerosas
cosas pueden pasar antes de ese límite de tiempo, una es que el electrón emita
el fotón reflejando la luz, pero otra muy diferente ocurre en la clorofila.
Cuando la clorofila es sometida a la luz de forma aislada esta refleja los
fotones que absorbe tornándose fluorescente, emitiendo los fotones en una
longitud de onda inferior a la que la recibió “parte de la energía se pierde en
forma de calor”. Sin embargo, si el mismo experimento es llevado a cabo en una
cantidad de cloroplastos que contienen la misma cantidad de clorofila, apenas
si se nota una muy leve fluorescencia. Es decir, en tales casos la energía
lumínica no se pierde de nuevo al ambiente, por el contrario, es almacenada por
el cloroplasto y empleada para realizar sus procesos de síntesis de alimentos.
En procariotas tales como cianobacterias, la fotosíntesis tiene lugar en gránulos unidos a la membrana plasmática o en un sistema de membranas internas propio llamado tilacoide. El sitio de la fotosíntesis en eucariotas como las plantas verdes y las algas verdes es el cloroplasto, una organelo que discutimos en la unidad de La Célula. Al igual que la mitocondria, el cloroplasto tiene membranas internas y externas y un espacio intermembrana.

Figura 3. El cloroplasto posee una serie de membranas internas donde
se sitúan complejos de proteínas encargados de evitar que la energía de la
clorofila escape de regreso al ambiente.
Además, dentro del cloroplasto hay cuerpos llamados grana,
que consisten en pilas de membranas aplanadas como monedas llamadas discos
tilacoides, el nombre común a las membranas de las cianobacterias no es ninguna
coincidencia, pues ambas hacen lo mismo con las mismas enzimas, lo cual es una
evidencia de su relación evolutiva (De Clerck, Bogaert, & Leliaert, 2012; Gray & Archibald, 2012). Las granas están conectadas
por membranas llamadas láminas intergranales. Los discos tilacoides se forman
mediante el plegado de una tercera membrana dentro del cloroplasto. El
plegamiento de la membrana tilacoide crea dos espacios en el cloroplasto además
del espacio intermembrana. El estroma se encuentra dentro de la membrana
interna y fuera de la membrana tilacoide. Además del estroma, hay un espacio intertilacoidal.
La captura de la luz y la producción de oxígeno tienen lugar en los discos tilacoidales.
Las reacciones oscuras (también llamadas reacciones independientes de la luz),
en las que el
Está bien establecido que el evento primario en la
fotosíntesis es la absorción de luz por la clorofila. Los estados de alta
energía (estados excitados) de la clorofila son útiles en la fotosíntesis
porque la energía luminosa se puede pasar a lo largo de complejos enzimáticos y
se convierte en energía química en las reacciones de la luz. Hay dos tipos
principales de clorofila, la clorofila a y la clorofila b. Los eucariotas tales
como las plantas verdes y las algas verdes contienen clorofila a y clorofila b.
Los procariotas tales como las cianobacterias (antes llamadas algas
azules-verdes) contienen sólo clorofila a. Las bacterias fotosintéticas
distintas de las cianobacterias tienen bacterioclorofilas, siendo la
bacterioclorofila a la más común. Los organismos como las bacterias verdes y
sulfuropurpurosas, que contienen bacterioclorofilas, no utilizan el agua como
la fuente última de electrones para las reacciones redox de la fotosíntesis, ni
producen el oxígeno. En su lugar, utilizan otras fuentes de electrones como
Los pigmentos son compuestos que tiñen de determinado color una solución, debido a que absorben y reflejan determinadas longitudes de onda. La luz absorbida son los colores que desaparecen de nuestra vista, mientras que la luz reflejada son los colores que aparecen a nuestra vista.

Figura 4. En el presente
espectro de absorción se muestra en el eje (x) las longitudes de onda de la
radiación electromagnética asociada a determinados colores. En el eje (y) se
encuentra el nivel de absorción relativo de determinada longitud de onda. La
clorofila a absorbe principalmente dos colores, el azul oscuro y el rojo
intenso, los demás colores los emite.
La clorofila que significa afín o amante del verde es por lo
tanto un nombre paradójico para los pigmentos verdes. Esto se debe a que los
pigmentos verdes absorben todos los colores, menos el verde el cual expulsan o
emiten al ambiente. Las hojas y una amplia gama de estructuras vegetales son
verdes debido a que sus cloroplastos poseen grandes cantidades de clorofila que
constantemente se encuentra emitiendo luz de longitud de onda verde que no es
absorbida para realizar la fotosíntesis. Los colores cuyas longitudes de onda
son más fuertemente absorbidos por la clorofila son el azul y el rojo.
Existen dos clases de clorofila, la clorofila a y la
clorofila b, ambos son pigmentos verdes que absorben longitudes de onda
especialmente en el azul y el rojo Figura
4.
En la Figura 5
podemos ver la estructura general de la clorofila, en verde el anillo de
porfirina con su centro de magnesio

Figura 5. Estructura general de la clorofila.
Las diferencias mínimas en la estructura, alteran las
longitudes de onda a las que responde cada molécula de clorofila. El resto de
la estructura es similar. La clorofila a es una molécula de dos partes.
La primera parte es un anillo de porfirina, que funciona como la parte de absorción de luz (Figura 5 en verde). Este anillo se encuentra en el interior de la membrana celular. La estructura recuerda a la del grupo heme, siendo un marco orgánico que apoya la función de un metal pesado. En la hemoglobina es un átomo de hierro, mientras que en la porfirina es un átomo de magnesio. El anillo de porfirina posee una propiedad similar a la del anillo del benceno, sus enlaces dobles y simples alternantes no son más que una representación del enlace deslocalizado en donde los electrones fluyen de manera poco unida a sus respectivos núcleos. Cuando un fotón de luz impacta contra el anillo de porfirina, se causa una redistribución de la densidad de electrones, los cuales, al estar deslocalizados, se liberan del anillo confacilidad. En otras palabras, la forma en como está unido el anillo de porfirina hace que los electrones de los enlaces dobles y simples sean fáciles de arrancar, y solo basta la energía contenida en un fotón de luz para lograr ese cometido. El anillo de porfirina vibra con los fotones de algunas longitudes de onda, mientras que otras simplemente pasan sin activar el sistema de emisión de electrones, siendo rebotados y generando su color, en otras palabras, el color de la clorofila es aquel que esta no emplea para la fotosíntesis.

Figura 6. Espectros de absorción de las clorofilas a y b,
en ambos casos los colores verdes no son absorbidos, lo cual hace que estas
sustancias se vean de ese mismo color.
Este detalle es particularmente evidente en un espectro de
absorción de absorción de luz para la molécula de clorofila (Figura 4
y Figura 6).
Al igual que cuando se hace para la fase gaseosa de ciertos elementos
purificados, el espectro de absorción es único, y permite identificar la
molécula. Sin embargo, ciertos polipéptidos cortos unidos de manera débil a la
clorofila pueden modificar el espectro de absorción de luz. Diferencias en la
absorción de luz entre las clorofilas a y b. Aunque ambas tienen picos de
absorción hacia los mismos colores "azul y rojo" no se sobrelapan
completamente, permitiendo absorber más gamas de ambos colores. Cabe anotar que
otra diferencia entre ambas clorofilas en las plantas terrestres es que se
asocian a un fotosistema específico.
A parte de la clorofila y los carotenoides existen otros pigmentos fotosintéticos que pueden absorber los verdes como la ficoeritrina y la ficocianina presente en las cianobacterias.

Figura 7. Espectros de absorción de
las clorofilas a y b, así como otros pigmentos como los carotenoides, la
ficoeritrina y la ficocianina, los cuales tienen absorciones en el verde, por
lo que se verán azules, anaranjados o rojos.
Muchas clases de clorofila no son reconocidas como tales
debido a una diversidad de polipéptidos accesorios que alteran las longitudes
de onda a las cuales responden. Las clorofilas sin alterar se encuentran más
ampliamente distribuidas en cianobacterias, algas y plantas terrestres, sin
embargo, las plantas terrestres tienen otras clorofilas modificadas. Esto se
debe a que el agua también es una molécula que responde a la luz, absorbiendo
longitudes de onda que se relacionan con colores que se pierden cuando la luz
ingresa al agua. En tierra esas longitudes de onda se mantienen y portan
energía. Estas clorofilas modificadas se denominan carotenoides, las cuales se
caracterizan por emitir luz no absorbida en los colores amarillo y anaranjado.
Cuando los tejidos vegetales acumulan una gran cantidad de
carotenoides adquieren colores característicos Amarillo y anaranjado como en la
zanahoria y la naranja. Estos pigmentos son acumulables cuando son consumidos
en el tejido adiposo de los mamíferos. Una persona con una dieta excesiva de
carotenoides se pondrá amarillo o anaranjado como una zanahoria. Los
carotenoides poseen funciones múltiples, en primera instancia en honor a su
origen filogenético, actúan como pigmento fotosintético secundario.
Adicionalmente ayudan a estabilizar a las clorofilas
extrayendo exceso de energía desde las clorofilas verdes y emitiéndola al ambiente
en forma de calor. Esto se debe principalmente a que el calor puede romper el
anillo de porfirina y otros componentes claves del sistema fotosintético. Otro
efecto perjudicial, es que el oxígeno absorbería esa energía calórica
haciéndolo hiperreactivo, y el oxígeno hiperreactivo es hipervenenoso,
hipermutagénico, hipercancerigeno, e hipermortal.
En 1932, Robert Emerson y William Arnold del Instituto Tecnológico de California llevaron a cabo una serie de experimentos que conllevaron a una extraña conclusión (Emerson & Arnold, 1932a, 1932b). No toda la clorofila en una planta era empleada para la conversión de energía lumínica en energía química. Empleando suspensiones del alga verde denominada Chlorella spp., y rayos de luz de duración extremadamente corta se pudo determinar la cantidad de energía mínima para la producción óptima de oxigeno durante la fotosíntesis. Basados en la cantidad de clorofila presente en la preparación, calcularon que una sola molécula de oxigeno era liberada durante un rayo de luz muy corto por cada 2500 moléculas de clorofila presentes. Posteriormente, Emerson demostró que se necesitaba un mínimo de 8 fotones para poder sintetizar una sola molécula de oxigeno molecular, lo cual implica que los cloroplastos poseen cerca de 300 veces las moléculas que deberían ser necesarias para oxidar el agua y sintetizar el oxígeno molecular. Lo anterior nos lleva a la pregunta ¿para qué sirve la clorofila extra?

Figura 8. La unidad fotosintética
actúa como una celda solar. Los carotenoides filtran la energía que dañaría a
la célula, mientras que las clorofilas transfieren la energía restante de
fotones a electrones vibratorios.
Una posible interpretación de estos resultados es que solo un pequeño porcentaje de las moléculas de clorofila están relacionadas con la fotosíntesis en sí misma, lo cual es una verdad a medias y una falsedad a medias. Las clorofilas se encuentran empaquetadas y actúan en grupos como uno solo, en una estructurada denominada unidad fotosintética. La unidad fotosintética posee clorofilas que se distribuyen las funciones, solo una parte de las clorofilas se desempeñan como el centro de reacción de la clorofila. La función del centro de reacción es transferir electrones a un aceptor. A pesar de que la mayor parte de las clorofilas no participan de manera directa en la conversión de luz en energía química, ellas son responsables de la absorción de la luz.

Figura 9. La unidad fotosintética se
divide en dos partes, la antena que transmite la energía y el centro de
reacción.
Las clorofilas y los carotenoides se los pueden clasificar
dos grupos diferentes, la antena y el centro de la reacción. En la antena se
encuentra la mayor densidad molecular de clorofilas, así como los carotenoides
que la protegen. La energía se transfiere desde los pigmentos que capturan
mayor energía a los que capturan menor energía. La transferencia de la energía
de excitación desde una molécula a otra es muy sensible a la distancia entre
las moléculas. Las moléculas en una antena de fotosíntesis se mantienen a una
gran proximidad unas de otras para no disipar la energía capturada.
Una regla que opera en los pigmentos de la antena
fotosintética, es que la energía solo se puede transmitir a una molécula que
pueda capturar la misma o menor energía. En otras palabras, la energía solo
puede pasar a una molécula que absorbe luz en la misma o en una longitud de
onda más grande “corriendo hacia el rojo”. A medida que la energía pasa de una
molécula a otra parte de ella se disipa y una fracción llega al centro de reacción,
la cual procesa menor energía que cualquier otra molécula ubicada en la antena.
Una vez el centro de reacción captura energía, este emite un electrón hacia su
aceptor.




No hay comentarios:
Publicar un comentario