
(Ciencias de Joseleg)(Biología)(Introducción y biología celular)(Los organelos celulares)(Introducción)(El citosol)(Organelos procariotas)(Los retículos endoplasmáticos)(El aparato de Golgi)(El núcleo celular)(Vesículas celulares)(La mitocondria y el cloroplasto)(Referencias bibliográficas)(Versión documento word)
El núcleo celular es el organelo que les da el nombre a las células eucarióticas. El nombre eucariota significa en el contexto científico, células con núcleos verdaderos. La función del núcleo celular es almacenar el genoma. Esto las diferencias de las células procariotas en las que no puede detectarse una membrana interna especializada en almacenar el genoma.

Figura 20. El núcleo es un organelo especial, pues su membrana es doble, y
parte de ella constituye al propio RER, además posee grandes poros.
Es por esto que células procariotas con sistema de membranas
internas como las cianobacterias se clasifican como procariotas, pues en el
caso de las cianobacterias ningún tilacoide almacena su genoma. El núcleo
celular está compuesto por dos membranas celulares, una interna y otra externa.
Ambas membranas se unen en regiones llamados poros. Es por esto que es posible
afirmar que el retículo endoplasmático rugoso hace parte estructural de la
membrana del núcleo. La membrane externa del núcleo celular es continua con la
membrana del retículo endoplasmático, sin embargo, aunque están unidos
físicamente cada una de las membranas poseen un diferente contenido de
proteínas, lo cual diferencia su funcionamiento.
Al interior de la membrana interna se encuentra el nucleoplasma el
cual contiene al genoma eucariótico. Cuando la célula se divide una región del
núcleo se tiñe densamente y se llama nucleolo, su función primordial es la de
ensamblar nuevos ribosomas. La función del núcleo es almacenar el genoma,
aislándolo del resto de la célula. El núcleo solo existe cuando la célula está
trabajando, es decir en las fases de crecimiento “G1 y G2” y en la fase de
síntesis “S” del ciclo celular. En su interior NO se almacenan los cromosomas,
lo que se almacena es la cromatina ya sea para replicar el ADN que contiene o
para generar ARNm. Debido a que el núcleo contiene la mayor parte del genoma de
la célula, produce mucho ARNm que es enviad a través de los poros nucleares
hacia el retículo endoplasmático rugoso.
El núcleo es el orgánulo más grande en las células animales.
En las células de mamíferos, el diámetro promedio del núcleo es de
aproximadamente 6 micrómetros (µm), que ocupa aproximadamente el 10% del
volumen celular total. El contenido del núcleo se mantiene en el nucleoplasma de
manera similar al citoplasma en el resto de la célula. El componente fluido de
esto se denomina nucleosol, similar al citosol en el citoplasma. En la mayoría
de los tipos de granulocitos, un glóbulo blanco, el núcleo está lobulado y
puede ser bilobulado, trilobulado o multilobulado.
La envoltura nuclear, también conocida como membrana nuclear, consta de dos membranas celulares, una membrana interna y otra externa, dispuestas paralelas entre sí y separadas por 10 a 50 nanómetros (nm). La envoltura nuclear encierra completamente el núcleo y separa el material genético de la célula del citoplasma circundante, sirviendo como barrera para evitar que las macromoléculas se difundan libremente entre el nucleoplasma y el citoplasma. La membrana nuclear externa es continua con la membrana del retículo endoplásmico rugoso (RER), y está tachonada de manera similar con ribosomas. El espacio entre las membranas se llama espacio perinuclear y es continuo con la luz RER.

Figura 21. La membrana nuclear es una estructura semicontínua con el
retículo endoplasmático rugoso.
Los poros nucleares, que proporcionan canales acuosos a través de la envoltura, están compuestos por múltiples proteínas, denominadas colectivamente nucleoporinas. Los poros tienen un peso molecular de aproximadamente 125 millones de daltons y consisten en alrededor de 50 (en levadura) a varios cientos de proteínas (en vertebrados). Los poros tienen un diámetro total de 100 nm; sin embargo, la brecha a través de la cual las moléculas se difunden libremente tiene solo unos 9 nm de ancho, debido a la presencia de sistemas reguladores dentro del centro del poro. Este tamaño permite selectivamente el paso de pequeñas moléculas solubles en agua al tiempo que evita que moléculas más grandes, como ácidos nucleicos y proteínas más grandes, entren o salgan inapropiadamente del núcleo. Estas moléculas grandes deben ser transportadas activamente al núcleo en su lugar. El núcleo de una célula de mamífero típica tendrá alrededor de 3000 a 4000 poros en toda su envoltura, cada uno de los cuales contiene una estructura en forma de anillo simétrica de ocho veces en una posición donde las membranas internas y externas se fusionan. Se une al anillo una estructura llamada canasta nuclear que se extiende hacia el nucleoplasma y una serie de extensiones filamentosas que llegan al citoplasma. Ambas estructuras sirven para mediar la unión a las proteínas de transporte nuclear.

Figura 22. Denominado complejo del poro nuclear, el poro es una estructura
compleja que media el paso de sustancias de manera selectiva.
La mayoría de las proteínas, las subunidades ribosómicas y
algunos ADN se transportan a través de los complejos de poros en un proceso
mediado por una familia de factores de transporte conocidos como carioferinas.
Esas carioferinas que median el movimiento hacia el núcleo también se llaman
importinas, mientras que las que median el movimiento fuera del núcleo se
llaman exportinas. La mayoría de las carioferinas interactúan directamente con
su carga, aunque algunas usan proteínas adaptadoras. Las hormonas esteroides
como el cortisol y la aldosterona, así como otras pequeñas moléculas solubles
en lípidos involucradas en la señalización intercelular, pueden difundirse a
través de la membrana celular y hacia el citoplasma, donde se unen a las
proteínas del receptor nuclear que son traficadas hacia el núcleo. Allí sirven
como factores de transcripción cuando se unen a su ligando; en ausencia de un ligando,
muchos de estos receptores funcionan como desacetilasas de histonas que reprimen
la expresión génica.
En las células animales, dos redes de filamentos intermedios
proporcionan al núcleo un soporte mecánico: la lámina nuclear forma una malla
organizada en la cara interna de la envoltura, mientras que se proporciona un
soporte menos organizado en la cara citosólica de la envoltura. Ambos sistemas
proporcionan soporte estructural para la envoltura nuclear y los sitios de
anclaje para cromosomas y poros nucleares.
La lámina nuclear está compuesta principalmente de proteínas laminares. Como todas las proteínas, las láminas se sintetizan en el citoplasma y luego se transportan al interior del núcleo, donde se ensamblan antes de incorporarse a la red existente de láminas nucleares. Las láminas que se encuentran en la cara citosólica de la membrana, como emerina y nesprina, se unen al citoesqueleto para proporcionar soporte estructural. Las láminas también se encuentran dentro del nucleoplasma, donde forman otra estructura regular, conocida como velo nucleoplasmático, que es visible mediante microscopía de fluorescencia. La función real del velo no está clara, aunque está excluida del nucleolo y está presente durante la interfase. Las estructuras laminares que forman el velo, como LEM3, se unen a la cromatina e interrumpen su estructura inhiben la transcripción de genes codificadores de proteínas.
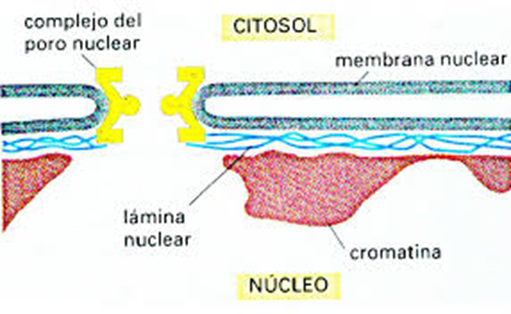
Figura 23. La lámina nuclear es un mediador proteínico entre la envoltura y
la cromatina.
Al igual que los componentes de otros filamentos
intermedios, el monómero lamina contiene un dominio alfa-helicoidal utilizado
por dos monómeros para enrollarse entre sí, formando una estructura de dímero
llamada bobina enrollada. Dos de estas estructuras de dímero se unen una al
lado de la otra, en una disposición antiparalela, para formar un tetrámero
llamado protofilamento. Ocho de estos protofilamentos forman una disposición
lateral que se tuerce para formar un filamento similar a una cuerda. Estos
filamentos pueden ensamblarse o desmontarse de manera dinámica, lo que
significa que los cambios en la longitud del filamento dependen de las tasas
competitivas de adición y eliminación de filamentos.
Las mutaciones en los genes laminares que conducen a
defectos en el ensamblaje de filamentos causan un grupo de trastornos genéticos
raros conocidos como laminopatías. La laminopatía más notable es la familia de
enfermedades conocidas como progeria, que causa la aparición de envejecimiento
prematuro en sus víctimas. El mecanismo exacto por el cual los cambios
bioquímicos asociados dan lugar al fenotipo envejecido no se conoce bien (Mounkes & Stewart, 2004).
El núcleo celular contiene la mayoría del material genético de la célula en forma de múltiples moléculas lineales de ADN organizadas en estructuras llamadas cromosomas. Cada célula humana contiene aproximadamente dos metros de ADN. Durante la mayor parte del ciclo celular, estos se organizan en un complejo de ADN-proteína conocido como cromatina, y durante la división celular se puede ver que la cromatina forma los cromosomas bien definidos familiares de un cariotipo. En cambio, una pequeña fracción de los genes de la célula se encuentra en las mitocondrias.

Figura 24. Los cromosomas son cromatina empacada y duplicada, se forman para
poder separar la cromatina durante la reproducción. Como el ADN está empacado,
se encuentra inactivo.
Hay dos tipos de cromatina. La eucromatina es la forma de
ADN menos compacta y contiene genes que la célula expresa con frecuencia. El
otro tipo, la heterocromatina, es la forma más compacta y contiene ADN que se
transcribe con poca frecuencia. Esta estructura se clasifica además en
heterocromatina facultativa, que consiste en genes que se organizan como
heterocromatina solo en ciertos tipos celulares o en ciertas etapas de
desarrollo, y heterocromatina constitutiva que consiste en componentes
estructurales cromosómicos como telómeros y centrómeros. Durante la interfase,
la cromatina se organiza en parches individuales discretos, llamados
territorios cromosómicos. Los genes activos, que generalmente se encuentran en
la región eucromática del cromosoma, tienden a ubicarse hacia el límite del
territorio del cromosoma.
Los anticuerpos contra ciertos tipos de organización de la
cromatina, en particular los nucleosomas, se han asociado con una serie de
enfermedades autoinmunes, como el lupus eritematoso sistémico. Estos se conocen
como anticuerpos antinucleares (ANA) y también se han observado junto con la
esclerosis múltiple como parte de la disfunción general del sistema
inmunitario. Como en el caso de la progeria, el papel desempeñado por los
anticuerpos en la inducción de los síntomas de las enfermedades autoinmunes no
es obvio (Zhang & Zhang, 2015).
El nucleolo es la más grande de las discretas estructuras sin
membrana, densamente teñidas, conocidas como cuerpos nucleares que se
encuentran en el núcleo. Se forma alrededor de repeticiones en tándem de ADNr,
ADN que codifica el ARN ribosómico (ARNr). Estas regiones se denominan regiones
organizadoras nucleolares (NOR). Los roles principales del nucleolo son
sintetizar rRNA y ensamblar ribosomas. La cohesión estructural del nucleolo
depende de su actividad, ya que el ensamblaje ribosómico en el nucleolo da como
resultado la asociación transitoria de los componentes nucleolares, facilitando
el ensamblaje ribosómico adicional y, por lo tanto, la asociación adicional.
Este modelo está respaldado por observaciones de que la inactivación del ADNr
resulta en la mezcla de estructuras nucleolares.
En el primer paso del ensamblaje de ribosomas, una proteína llamada ARN polimerasa I transcribe ADNr, que forma un gran precursor de ARNt previo. Esto se divide en las subunidades 5.8S, 18S y 28S rRNA. La transcripción, el procesamiento postranscripcional y el ensamblaje del rRNA se produce en el nucleolo, ayudado por pequeñas moléculas de ARN nucleolar (snoRNA), algunas de las cuales se derivan de intrones empalmados de ARN mensajeros que codifican genes relacionados con la función ribosómica. Las subunidades ribosómicas ensambladas son las estructuras más grandes que pasan a través de los poros nucleares.

Figura 25. El nucléolo es el punto de ensamblaje de los ribosomas.
Cuando se observa bajo el microscopio electrónico, se puede
ver que el nucleolo consta de tres regiones distinguibles: los centros
fibrilares más internos (FC), rodeados por el componente fibrilar denso (DFC)
(que contiene fibrillarina y nucleolina), que a su vez está bordeado por el
componente granular (GC) (que contiene la proteína nucleofosmina). La
transcripción del ADNr se produce en el FC o en el límite de FC-DFC y, por lo
tanto, cuando aumenta la transcripción del ADNr en la célula, se detectan más
FC. La mayor parte de la escisión y modificación de los rRNA se produce en el
DFC, mientras que los últimos pasos que implican el ensamblaje de proteínas en
las subunidades ribosómicas se producen en el GC.
Además del nucleolo, el núcleo contiene varios otros cuerpos
nucleares. Estos incluyen los cuerpos de Cajal, los géminis de los cuerpos de
Cajal, la asociación de cariosomas de interfase polimórfica (PIKA), los cuerpos
de leucemia promielocítica (LMP), los parapeckle y las manchas de empalme.
Aunque se sabe poco sobre varios de estos dominios, son significativos porque
muestran que el nucleoplasma no es una mezcla uniforme, sino que contiene
subdominios funcionales organizados.
Otras estructuras subnucleares aparecen como parte de
procesos anormales de la enfermedad. Por ejemplo, se ha informado de la
presencia de pequeñas varillas intranucleares en algunos casos de miopatía por
nemalina. Esta condición generalmente es el resultado de mutaciones en la
actina, y las barras en sí consisten en actina mutante, así como otras proteínas
del citoesqueleto (Mao, Zhang, & Spector, 2011).




{kind=link}
No hay comentarios:
Publicar un comentario