(Ciencias de Joseleg)(Biología)(Introducción y biología
celular)(La célula)(Introducción)(Del
vitalismo a la abiogénesis)(El
fin del vitalismo)(Del
microscopio a la teoría celular)(Más
que una bolsa de proteínas)(Como
se estudia la célula)(Las
propiedades de las células)(Generalidades
de la célula y su estudio)(La
célula procariota)(La
célula eucariota)(Introducción
a las partes de la célula)(Referencias
bibliográficas)(Versión
documento word)
Protistos
En muchos aspectos, las células eucariotas más complejas y
capaces no se encuentran en el interior de las plantas y los animales. Estas
células se encuentran en el interior de los muchos grupos de seres vivos
unicelulares eucarióticos llamados protistos. Los protistos son un enorme grupo
de seres vivos, los cuales en términos morfológicos aunque están compuestos por
células eucariotas, no se los podía clasificar ni como un hongo, una planta o
un animal, ellos son sus propios tipos de seres vivos eucariotas, tan
diferentes a un animal, como lo es una planta a un hongo.

Figura 41. Protistas como Dictyostelium spp., se encuentran en la
transición entre la unicelular y lo multicelular (YouTube).
Giardia lamblia expone
la diversidad celular de los eucariotas, un ser vivo que mantiene la mayor
parte de su ciclo con dos núcleos haploides independientes y activos. El estado
de dicarionte como se describe a tener dos núcleos que no se unen también se
puede ver en los hongos y de hecho brevemente en el cigoto humano. Entre los
protistas unicelulares más comúnmente conocidos contamos a las amebas, las
euglenas y el volvox. Estas células se caracterizan por realizar todas las
funciones vitales, a diferencia de los eucariotes multicelulares donde se da la
tendencia a la especialización. Sin embargo, existen casos en los que un
protista puede presentar un estado de vida unicelular, pero si las condiciones
cambian pueden pasar a ser multicelulares con una especialización celular
relativamente poco avanzada. Los protistas no son solo unicelulares, si no
también multicelulares, tal como Dictyostelium
spp., un ser vivo que en vida libre se comporta como una ameba, pero
cuando las condiciones se hacen duras se congrega sobre sí mismo formando
pequeñas babosas y luego una estructura fructificante, Sus células en la estructura
fructificante experimentan diferenciación, donde las de la punta realizan la
reproducción sexual, mientras que las del talo se sacrifican para que la
población sobreviva en otro lugar.
Una forma alternativa de la evolución en el modo de vida individualista, fue el modo de vida grupal, en donde la célula individual pierde importancia en favor de la supervivencia del grupo. Los modos de multicelularidad no son igualmente complejos. Como ya mencionamos anteriormente, existen protistas generalmente unicelulares que se agrupan en ciertas eventualidades ambientales generando niveles de especialización celular parcial en donde solo algunos individuos lograran reproducirse, mientras que el resto se muere y no se reproduce, sacrificándose en favor del futuro.

Figura 42. La
multicelularidad ha evolucionado varias veces, una de las más recientes se ha
dado en el linaje del Volvox, un alga
verde microscópica de la cual se tienen los estadios morfológicos de varios
puntos. Por ejemplo, dentro de Volvox tenemos las Chlamidiomonas
que son incelulares de vida libre, luego tenemos a Gonium que es una
agregación celular. Luego tenemos a Eudoria que ya posee una matriz
extracelular, lo que mantiene a las células unidas pero a cierta distancia unas
de otras. En Pleodorina ya tenemos un nivel incipiente de
especialización celular, mientras que Volvox como tal posee tejidos
tan especializados como el germinal.
Las esponjas también son agregados celulares, las células
presentan una pobre especialización y los tipos de células apenas si pueden ser
contados con los dedos de las manos, mientras que los tipos de células de un
ser humano pueden llegar a más de 200. En los seres multicelulares donde la
multicelularidad se va acentuando, el estado multicelular se hace más
prolongado, y se hace el estado en que la especie se alimenta. En estos seres
vivos, las células de la comunidad sufren especializaciones muchísimo más
marcadas. Esta especialización celular es llevada a cabo mediante el proceso de
diferenciación celular durante el desarrollo embrionario de la especie.
Si las esponjas son simples los placozoos lo son aún más, literalmente son una masa de células con flagelos que le permite el movimiento, ostenta el título de animal más simple y probablemente sea un ser vivo simplificado a partir de otro con verdaderos tejidos. A diferencia de lo ocurre con los seres vivos como Dicyostelium spp., la diferenciación celular en los eucariotes multicelulares es mucho más marcada, un solo embrión humano puede dar lugar a 200 tipos de células diferentes, con formas, funciones y bioquímica completamente diferentes, las cuales se coordinan para generar un todo, una unidad coordinada, un ser vivo multicelular.
Como resultado de la diferenciación, diferentes tipos de
células adquieren una apariencia distintiva y contienen materiales únicos,
llevan a cabo reacciones químicas únicas y se encuentran en zonas únicas de la
anatomía del ser vivo. El cigoto puede reconocerse porque es un dicarionte, a
diferencia de lo que se esperaría, los núcleos del ovulo y el espermatozoide no
experimentan la cariogamia, en lugar de ello, iniciarla primer mitosis o
clivaje, y en el momento en que las membranas de los dos núcleos se disuelven,
el material genético se duplica y se reorganiza de modo que las dos células del
embrión si poseen el estado diploide, pero nunca se fusionan los núcleos. A
partir del cigoto se generan cientos de tipos de células diferentes.
En los mamíferos por ejemplo tenemos las células
esqueléticas, musculares, glandulares, nerviosas, epiteliales, sanguíneas,
protectoras entre muchas otras más. Cuando el embrión alcanza el nivel de
gástrula, las células que lo componen ya se han diferenciado en tres tipos
generales, células del ectodermo, del mesodermo y del endodermo. Cada una de
estas tres dará lugar a un tipo de tejido y de órganos diferentes.
Una pregunta pertinente seria, ¿Cómo se lleva a cabo la
diferenciación celular? Anteriormente se pensaba que las células eliminaban los
genes innecesarios a medida que se iban diferenciando unas de otras. Mientras
que actualmente que más que eliminar, los genes se van activando y
desactivando. Prueba de esto es que, si se realiza una transferencia de un
núcleo somático a un ovulo, el ovulo activa nuevamente todos los genes del
núcleo reiniciando el programa y generando un embrión clonado de manera
somática.
Las células eucariotas tienen una cantidad limitada de rutas metabólicas, pero en compensación, poseen una gama casi infinita de formas especializadas, a sea en los organismos de vida libre, parásitos, o en organismos multicelulares, sin embargo, para simplificar las cosas, por tradición se introducen solo dos, una célula animal idealizada, y una célula vegetal idealizada, pero el lector deberá tener en cuenta que existen muchos tipos y formas diferentes de las presentadas en este documento.

Figura 43. Estructura idealizada de la célula animal.
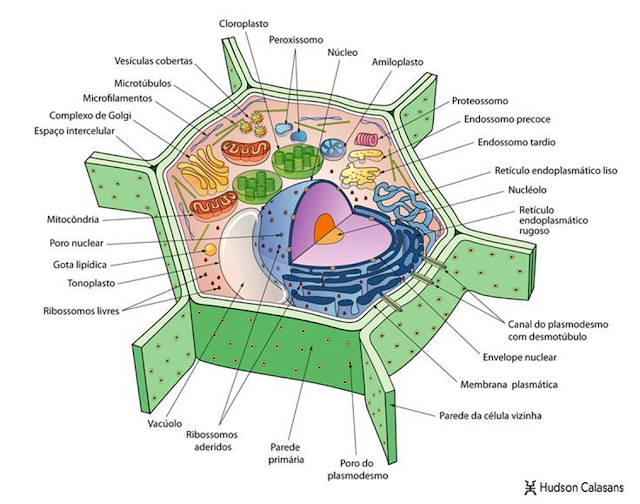
Figura 44. Estructura idealizada de la célula vegetal.

Figura 45. Toxoplasma gondii es un parásito intracelular que aprovecha el ciclo depredador presa, sus células son de tipo eucariota al tener núcleo y un sistema de membranas internas, pero adicionalmente posee organelos especializados que no son referenciados en los cursos introductorios de biología celular.
Deberías notar que las células eucariotas están altamente compartimentadas. Estos compartimentos están formados por membranas que crean espacios internos que dividen el trabajo necesario para llevar a cabo las funciones de la vida. Los compartimentos de una célula eucariota, típicamente llamados organelos u orgánulos, llevan a cabo funciones especializadas que juntas permiten que la célula sea más eficiente y exitosa. Casi todos los orgánulos están rodeados por una membrana con proteínas integradas, muchas de las cuales son enzimas (moléculas que aceleran las reacciones químicas). Estas enzimas hacen productos específicos para ese orgánulo, pero su acción beneficia al sistema celular completo.
La célula se puede ver como un sistema de organelos
interconectados que trabajan juntos para conducir y regular los procesos de la
vida. Por ejemplo, el núcleo es un compartimento que alberga el material
genético dentro de los cromosomas eucariotas, que contienen información
hereditaria. El núcleo se comunica con los ribosomas en el citoplasma, y los
orgánulos del sistema de endomembranas, especialmente el retículo endoplásmico
(RE) y el aparato de Golgi, se comunican entre sí.
La producción de moléculas específicas tiene lugar dentro o
sobre la superficie de los orgánulos. Estos productos son luego transportados
alrededor de la célula por vesículas de transporte, sacos membranosos que
encierran las moléculas y las mantienen separadas del citoplasma. Por ejemplo,
el retículo endoplásmico se comunica con el aparato de Golgi por medio de
vesículas de transporte.
Las vesículas se mueven por medio de una extensa red de
fibras de proteína llamada citoesqueleto, que también mantiene la forma de la
célula y ayuda con el movimiento de la célula. Estas fibras de proteínas permiten
que las vesículas se muevan de un orgánulo a otro. Los organelos también se
mueven de un lugar a otro usando este sistema de transporte. Piense en el
citoesqueleto como un sistema de vías tridimensional dentro de las celdas
utilizadas para transportar carga importante de un lugar a otro. Los orgánulos
relacionados con la energía -cloroplastos en plantas y mitocondrias, tanto en
plantas como en células animales- son responsables de generar la mayor parte de
la energía necesaria para realizar procesos celulares.